Introduction
The sessile nature of plants has led to unique adaptations to their environment. Throughout evolution, plants have survived by changing their functional or structural characteristics. Animals, being mobile, can change their circumstances to some extent, but plants cannot. However, plants are not static; they can respond to a variety of environmental stimuli. Plants perform various macroscopic and microscopic movements in response to intrinsic and extrinsic stimuli.
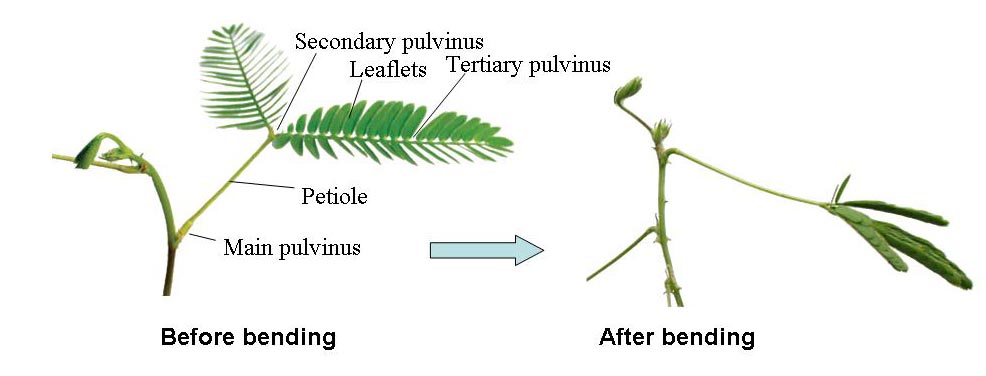
Mimosa pudica L. responds to mechanical, thermal, electrical, and chemical stimuli by rapidly bending leaves and petioles. Like other members of the mimosa family, the leaves of M. pudica close at night or in the dark and open during the day or in the light. Thus, the sensitive M. pudica has the ability to perform both seismonastic and nyctinastic movements. The movement of leaves and petioles in Mimosa is controlled by changes in the curvature of the pulvinus. Leaf folding and petiole bending are caused by rapid shrinkage of the underside of the motor cells of the pulvinus. Stimulation of a leaf causes an electrical signal to propagate up the petiole. Subsequently, the electrical signal is converted into a chemical signal that causes certain cell membranes to become more permeable to K+ and Cl- ions. The increased permeability of the cell membranes causes ions to move from the symplast to the apoplast. Water accompanies the ions into the apoplast by osmosis. This phenomenon is called plasmolysis. As a result of plasmolysis, the protoplast shrinks, causing the leaflets or petioles to droop.
REGULATORY MECHANISM
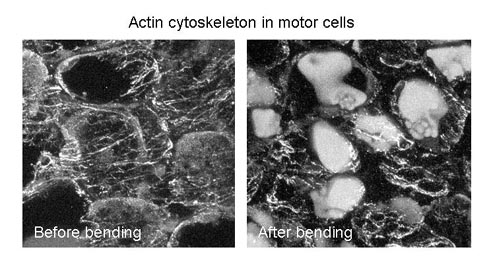
Our research focused on understanding how the cell's structural framework, known as the cell cytoskeleton, contributes to the unique movement of Mimosa plants when they respond to stimuli like touch. The cytoskeleton, composed of proteins like actin, serves as a kind of "scaffolding" inside cells, crucial for various cell functions.
To investigate this, we treated the part of the Mimosa plant responsible for movement with substances that disrupt actin, such as cytochalasin B and phalloidin. These substances essentially mess with the actin structure. What we observed was that when we interfered with actin, Mimosa lost its ability to bend in response to stimuli. This suggests that actin rearrangement is essential for the plant's movement.
Furthermore, we discovered that when the actin filaments were fragmented and certain acidic actin of four isoforms were reduced, Mimosa's bending response was affected. Interestingly, we found that inhibitors of a specific enzyme called Tyr phosphatase could prevent this effect. This indicates that there's a continuous process involving the addition and removal of chemical groups to actin, and this cycle is crucial for Mimosa's movement.
In essence, our study reveals that the structure of actin inside cells is vital for Mimosa's unique ability to move when touched. By understanding these molecular processes, we gain insights into how plants respond to their environment, which could have broader implications for plant biology and even applications in fields like agriculture.
ACTIN BINDING PROTEINS
Recent investigations have focused on molecular mechanisms regulating the actin cytoskeleton in the motor cells. In animals, formation and stability of actin filaments is controlled by actin-binding proteins that affect actin nucleation, actin monomer sequestration, and actin filament (F-actin) severing, capping, or bundling. Several actin-binding proteins have also been identified biochemically and molecular biologically in plants. They are thought to be responsible for modulating changes in actin organization and the dynamics of plant cell morphogenesis and development. We have shown that the presence of a protein similar to the gelsolin family proteins that severs actin filaments in a Ca2+-dependent manner. We also isolated annexin from the plant. Plant annexins are distributed in a wide variety of tissues and are able to bind actin in a calcium- or pH-dependent manner. Mimosa annexin is localized to the plasma membrane of pulvinar cells during the daytime. However, we observed a drastic change in distribution at night. Thus, Mimosa annexin appears to contribute to membrane reorganization during nyctinastic movements. In higher plants, villin, which was originally isolated from the actin bundles of intestinal epithelial cell microvilli, promotes formation of stable actin cables in vivo and in vitro (Yokota & Shimmen). Our present observations suggest that an actin-bundling protein is involved in the formation of actin cables, and the activity of such a protein appears to be regulated in response to external stimuli. So far, we focus on the contribution of villin in the seismonastic movement.
AQUAPORINS
As I mentioned above, an increased permeability causes water and ions to move from the symplasts into the apoplasts. Over the course of a few seconds, water in the extensor half of the pulvinus is transferred to the flexor half, resulting in movement of the leaf. Thus, water tansport is a key factor for the initiation of the seismonastic movement. Aquaporins (AQPs) are water channels that influence changes in turgor pressure in plant cells by transporting water across cell membranes. We isolated cDNAs encoding Mimosa aquaporins and expressed these clones in Xenopus oocytes. Mimosa aquaporins PIP2;1 and TIP1;1 show high water channel activity. When both clones were coexpressed in Xenopus oocytes, the water channel activity of the oocytes were increased rather than that of single expression. We also found that only PIP1;1 was phosphorylated by PKA at Serine131, and this phosphorylation enhanced the cooperative regulation of PIP isoforms. Thus, we believe that the permeability of membranes of Mimosa to water is regulated by both direct interactions and phosphorylation of AQP isoforms.
